What does sex really cost?
Identifying these costs is a major challenge. It has been said by some that theoretical work can exclude benefits of sex and simply quantify the cost as the fitness ratio of asexual to sexual organisms. However, this does not completely work. If immediate benefits of sex exist, the fitness ratio will underestimate the cost aspect of the equation (Lehtonen et al., 2012). Asexual counterparts might not even exist or be suitable for comparing for a given sexual species. Therefore, it must be kept in mind that researchers should be aware of taxonomic variation in life histories and reproductive modes. This includes for example whether the species in question is unicellular or multicellular, hermaphroditic or with distinct males and females.
Two-fold Cost of Sex:
One of the most talked about cost of sex is the "two-fold cost of sex". During the 1970s, George Williams and John Maynard Smith both produced theories trying to explain the two-fold cost of sex. These two theories spoke about the cost linking to genome dilution and male production, respectively. Genome dilution refers to all the genes of asexually produced offspring being from their single parent, whereas only half the genes in the offspring of a sexual female are from the mother.The representation of maternal genes is reduced by 50% each generation. From this it has been inferred that unless a sexual parent produces twice as many offspring per individual, they suffer a transmission disadvantage - this problem was labelled "the cost of sex" (Bell 1982). Whereas, the cost of males links in many species but not all, to the issue that males do not provide any resource to the next generation, yet sexual females typically invest half of their resources into the production of males. Everything else being equal, this generates a 2-fold advantage for asexual females (producing only female offspring). Another cost of sex is breaking favourable genetic combinations. While sexual reproduction enhances the potential for evolution, this is due to recombination as it expands the range of gene combinations. However, in the short term, recombination is costly because it breaks up successful gene combinations. The success of particular gene combinations relies on non-additive genetic interaction. This reshuffling of the sets of genes that worked well together in the previous generation, reflects the cost of sex owing to recombination as new genotypes in the next generation are, on average, less fit. Formally, the decline in the mean fitness of a trait owing to recombination is referred to as ‘genetic slippage’. Therefore, the more stable the environment, the greater the likelihood that reshuffled genotypes will be less fit than previously successful ones, increasing the cost of recombination (Lehtonen et al., 2012).
Mate Choice:
The evolution of mate choice has a cost before it even begins, as there is cost involved in finding a potential mate. Mate choice occurs for both male and females. This evolution of mate choice has led for some species, to produce exaggerated traits e.g. male peacock tail (Figure 1.) or behaviours e.g. male bowerbird (SciShow, 2016 from 1:15). While these displays are often filmed among bird species other organisms such as fish and spiders also show these traits e.g. Peacock Spider. However these exaggerated traits have the potential to be detrimental towards the individual for example, the male peacock tail needs a high energy input to produce and maintain, whilst also putting the male at a greater risk from predation. The "sexy son mechanism" (Fisher, 1930) can be estimated to aim us in deciding and explaining why male ornaments and displays are often extreme, and differ more than Figure 1. Female peacock observing a male peacock displaying. other traits between closely related species (Andersson et al., 2006). These traits apparently evolve rapidly due to the high selection pressure through mate choice in this case and have been linked in the involved in speciation.
to produce exaggerated traits e.g. male peacock tail (Figure 1.) or behaviours e.g. male bowerbird (SciShow, 2016 from 1:15). While these displays are often filmed among bird species other organisms such as fish and spiders also show these traits e.g. Peacock Spider. However these exaggerated traits have the potential to be detrimental towards the individual for example, the male peacock tail needs a high energy input to produce and maintain, whilst also putting the male at a greater risk from predation. The "sexy son mechanism" (Fisher, 1930) can be estimated to aim us in deciding and explaining why male ornaments and displays are often extreme, and differ more than Figure 1. Female peacock observing a male peacock displaying. other traits between closely related species (Andersson et al., 2006). These traits apparently evolve rapidly due to the high selection pressure through mate choice in this case and have been linked in the involved in speciation.
Predation:
Predation risk is a cost of sexual reproduction. Higher risk may be associated with all stages of reproduction. Examples of evolutionary responses to this increased risk include habitat choice, duration and timing of display and copulation, changes in brightness of breeding colouration, and changes in life history traits such as age of reproduction and reproductive effort.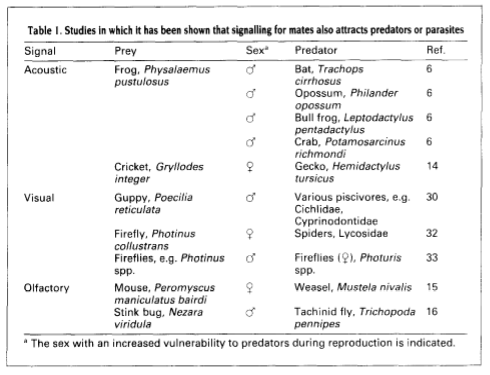
One of the main examples of increased risk includes signalling for mates, as the predators of the species signalling have now evolved awareness of the signal. Therefore, when the prey in question has been signalling a hopeful mate it also has the potential risk of attracting its predator to its location.
Table 1. (Left) Taken from (Magnhagen 1991)
Sexual Conflict:
Alongside arises sexual conflict, which could be between mates or be produced from the competition for mates. It appears that sexual conflict between males and females is the rule rather than the exception, sometimes leading to sexually antagonistic coevolution (Williams, 1966; Trivers, 1972; Parker, 1979). In terms of mate-choice evolution, sexual conflict can impose direct selection on the female preference that leads to increased female resistance to the male trait, because its consequences are negative for females. For example seed beetles (Coleoptera bruchidae). Spines developed on the male genitalia helps enhance stability when mating and thereby greater success in copulation (Edvarsson & Tregenza, 2005). These male seed beetles have also now been re corded showing the evolution of genital spines while female seed beetles developed tougher copulatory ducts to resist the spines (Arnqvist et al., 2007). Many aspects of sexually antagonistic coevolution, such as its co-occurrence with indirect selection on mate choice are still debated. There are also several more traits over which sexual conflict can occur, e.g. conflicts over limited resources, infanticide (Trivers, 1972; Hrdy, 1977) and mate cannibalism (Parker, 1979). Figure #. (Right) Female Argiope bruennichi and her much smaller mate whom is about to be consumed (Welke, 2012)
corded showing the evolution of genital spines while female seed beetles developed tougher copulatory ducts to resist the spines (Arnqvist et al., 2007). Many aspects of sexually antagonistic coevolution, such as its co-occurrence with indirect selection on mate choice are still debated. There are also several more traits over which sexual conflict can occur, e.g. conflicts over limited resources, infanticide (Trivers, 1972; Hrdy, 1977) and mate cannibalism (Parker, 1979). Figure #. (Right) Female Argiope bruennichi and her much smaller mate whom is about to be consumed (Welke, 2012)
Given these high costs of sex and the potential for asexual reproduction has led to a paradox . Most scientists might answer that sex and recombination have evolved as they generated variation needed by selection which links to one of the oldest explanations for sex "Sexual reproduction can also increase the differences between individuals... Such differences afford the material by means of which natural selection is able to increase or weaken each character according to the needs of the species." (Weismann, 1889)